EPIGENETIKA
Povratak Lamarckovih žirafa. Mogu li se stečene promjene prenijeti na buduće generacije?
 Tamara Čačev / 17. ožujka 2023. / Perspektive / čita se 16 minuta
Tamara Čačev / 17. ožujka 2023. / Perspektive / čita se 16 minuta

Group giraffe in National park of Kenya, Africa
Tamara Čačev / 17. ožujka 2023. / Perspektive / čita se 16 minuta
Group giraffe in National park of Kenya, Africa
Lamarckova ideja naslijeđivanja, po kojoj potomci nasljeđuju osobine koje su njihovi roditelji stekli za života, danas je gotovo u potpunosti zasjenjena Darwinovom idejom o prirodnoj selekciji iza koje stoji planina znanstvenih dokaza, piše Tamara Čačev. No, krajem prošlog stoljeća postalo je jasno da postoje i nasljedni elementi iznad genetičke osnove, što je ponovno na površinu izbacilo Lamarckova promišljanja.
Devetnaesto stoljeće je u mnogočemu bilo stoljeće uzleta ljudske misli i inovacija. U njemu su mnoga područja krenula znanstveno utemeljenim smjerovima. Pogotovo je to slučaj u području biologije jer je to stoljeće obilježeno istraživanjima Mendela u području klasične genetike te Darwinovih i Wallaceovih promišljanja evolucije. Istraživanja nasljedne osnove u dvadesetom stoljeću potvrdila su zakonitosti nasljeđivanja koje je postulirao Mendel, dok je molekularna filogenija potvrdila postavke Darwinove teorije evolucije. Ipak, valja reći kako u vrijeme Mendela i Darwina, koji su bili suvremenici, ništa od toga nije bilo poznato, nije se znala tvar koja predstavlja nasljednu osnovu o kojoj je govorio Mendel niti je bilo jasno na koju materijalnu osnovu ljudskog bića djeluje prirodna selekcija. Naravno, riječ je o molekuli DNA. Kao kuriozitet ostaje pitanje je li Darwin bio upoznat s radom Mendela, kako neki izvori tvrde, no čak i da jest, upitno je koliko bi u ono doba, s ondašnjim spoznajama mogao išta bitno unaprijediti u svojoj teoriji, obzirom na nepoznavanje materijalne osnove nasljeđivanja.
U promišljanju evolucije postoji jedno „kokoš ili jaje pitanje“ oko kojeg su se do pojave moderne biologije lomila koplja. Vjerojatno ste negdje načuli pojam Lamarckove žirafe. Naravno, nije se radilo o njegovom kućnom ljubimcu nego o primjeru koji je postao emblematičan za njegov rad, ponekad i za izrugivanje, iako zapravo taj primjer nije ni približno okosnica onog o čemu je Lamarck pisao. Radi se naime o jednom evolucijski zanimljivom pitanju: ima li žirafa dugačak vrat jer se morala propinjati do visokih grana da dođe do hrane pa se on stoga korištenjem produljio ili su u populaciji „protožirafa“ da ih tako nazovemo, postojale jedinke s duljim vratovima te su stoga bile odabrane prirodnom selekcijom da opstanu, dok su kratkovrate imale manje mogućnosti da dođu do hrane te su stoga otpale iz populacije jer su umrle ili bile nedovoljno uhranjene za prokreaciju. Prva verzija je ona koju je zastupao Lamarck, dok je drugu zagovarao Darwin.
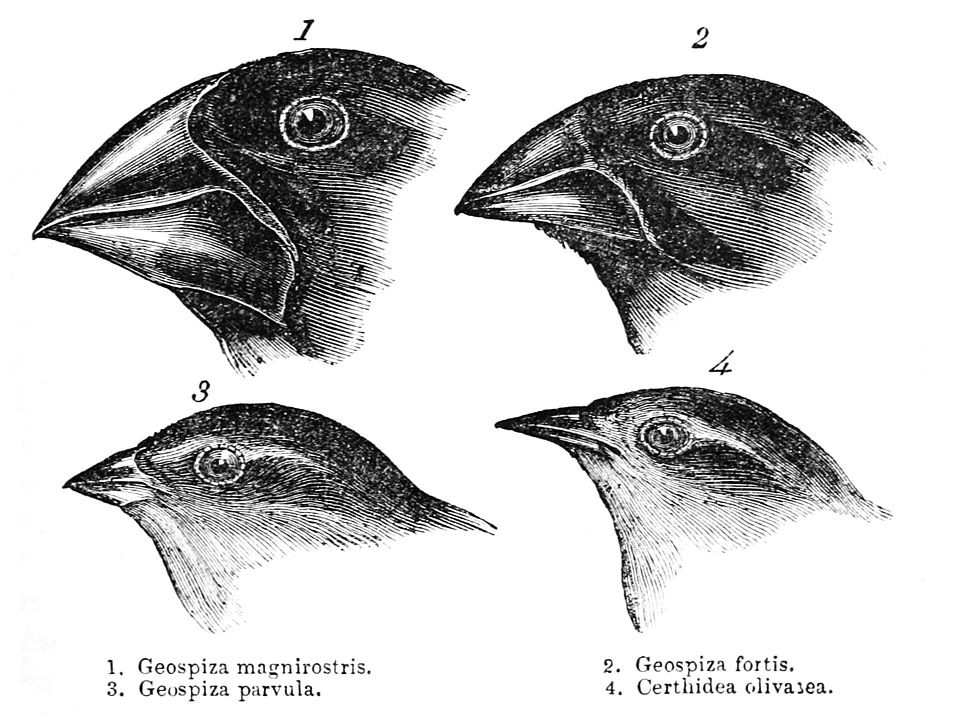
Danas je prihvaćeno stajalište temeljeno na Darwinovim promišljanjima da prirodna selekcija djeluje na varijabilnost fenotipa (svojstava koja primjećujemo kod nekog organizma) kroz genetičku varijabilnost koja je sama po sebi slijepa na utjecaje okoliša. Pojednostavljeno rečeno, varijabilnost u svojstvima živih bića je njima inherentna, to je svojevrsni švedski stol s kojeg će se iz cijele zbirke mogućih genetičkih varijanti u molekulama DNA prirodnom selekcijom odabrati one verzije genotipa koje u nekom trenutku pritiska čimbenika okoliša daju najbolje adaptiran fenotip. Da ponovimo, genotip je zapis u molekuli DNA a fenotip je njegova manifestacija poput boje kose ili očiju, ali i mnogih drugih, oku nevidljivih svojstava poput toga koliko brzo metaboliziramo neki lijek. Nasuprot ovako formuliranoj Darwinovoj teoriji evolucije dugo je bio suprotstavljen koncept koji je postavio Lamarck a temeljio se na ideji da se organizam modificira u skladu s uvjetima u okolini i onda ta stečena „poboljšanja“ prenosi na svoje potomstvo. Drugim riječima, okoliš može utjecati na svojstva organizma i zatim se ta „stečena“ svojstva mogu prenositi na iduću generaciju.
Međutim, istraživanjima Weismana i postuliranjem biološkog koncepta „germplasm“, prema kojem su tjelesne stanice strogo odvojene od spolnih, te da ne postoji način da utjecaj okoliša na tjelesne stanice ikada modificira svojstva spolnih stanica, smatralo se da je ideja lamarkizma zapravo pokopana. Nasljedna informacija ne može se prenijeti iz tjelesne stanice u spolnu i ta se prepreka naziva još i Weismannovom barijerom. Iz spolnih stanica nastaju tjelesne stanice nekog višestaničnog organizma, no štogod da te stanice steknu od svojstava tijekom života ne prenosi se na spolne stanice i dalje u iduću generaciju. Ovo pravilo je temelj suvremene genetike. Ipak, krajem dvadesetog stoljeća postalo je jasno da povrh genetičkih principa vezanih uz nasljednu osnovu tj. molekulu DNA postoji i područje genomske regulacije koje se nalazi „iznad“ (grčki. epi) te genetičke osnove te se stoga naziva epigenetičkom regulacijom, opet je na površinu izbacilo stara-nova promišljanja koja je predložio Lamarck.
Epigenetička regulacija ne odvija se na razini zapisa u molekuli DNA već se odnosi na razinu ekspresije gena koji su njome kodirani. Pod tim se stoga podrazumijevaju promjene u ekspresiji genoma koje ne uključuju izmjene na razini sekvence molekule DNA. Radi se o modifikacijama molekule DNA koje nisu mutacije već je riječ o metilaciji molekule DNA ili o modifikacijama proteina histona oko kojih se molekula DNA namata i tako pakira u kompaktniji oblik kromosoma.
I jedne i druge promjene ne mijenjaju zapis molekule DNA, no mijenjaju način kako se taj zapis interpretira, tj. kako se geni eksprimiraju. Ako to usporedimo s notnim zapisom, onda on i dalje ostaje isti, no oznakama za tempo možemo značajno utjecati na to kako neku glazbu interpretiramo jer adagio i allegro daju sasvim drugačiji dojam jednog te istog notnog zapisa. Iako, da budemo točniji, epigenetske modifikacije bi zapravo odgovarale promjenama tipa odsviraj ili preskoči neki stavak.
Etabliranje epigenetičke regulacije kao validnog mehanizma regulacije ekspresije gena ponovno je otvorilo vrata za razmatranje zasad još uvijek kontroverznoj ideji nasljeđivanja takvih epigenetičkih promjena a time potencijalno i osobina koje su stečene tijekom života, te je dovelo do sasvim novog koncepta koji se danas naziva epigenetičkim nasljeđivanjem. Ovaj se pojam odnosi na izmjene svojstava koja se prenose kroz generacije, a nisu rezultat promjena u samom zapisu u molekuli DNA. Kako bi se shvatilo o čemu se ovdje radi najprije treba jasno razdvojiti pojam intergeneracijskog od transgeneracijskog epigenetičkog nasljeđivanja. Dok se intergeneracijsko epigenetičko nasljeđivanje odnosi na prijenos epigenetičke informacije s roditelja na djecu, pojam transgeneracijskog nasljeđivanja odnosi se na prijenos epigenetičke informacije u generacije nakon ove prve, dakle na unuke i njihove potomke.
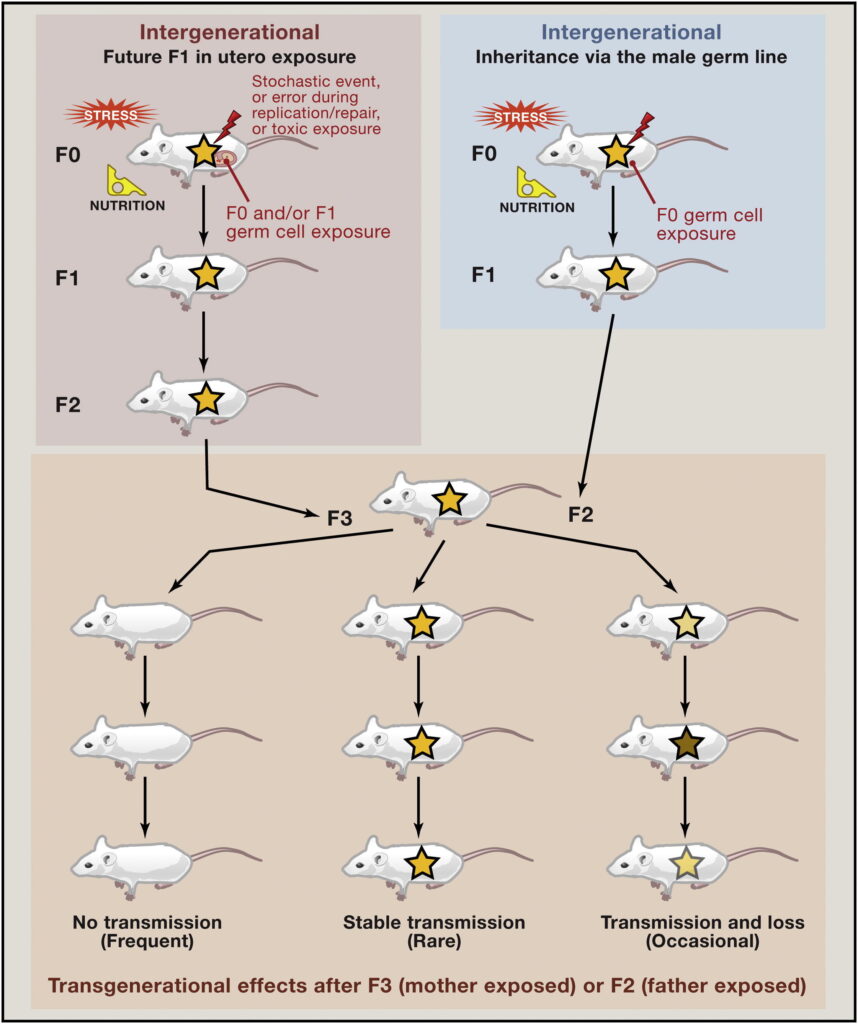
Kako bi to onda zapravo izgledalo na nekom plastičnom primjeru? Epigenetičke promjene u sisavaca mogu nastati uslijed izloženosti određenim utjecajima okoliša poput stresa, toksina, hormona ili nutritivnih deficita. Ako se radi o ženskoj trudnoj jedinki tada je in utero tom djelovanju izložen i embrij (F1 generacija) (tzv. parentalni, roditeljski ili intergeneracijski učinak) ali i njegove spolne stanice (koje će u konačnici rezultirati idućom F2 generacijom – unucima u odnosu na trudnu ženku). Takav intergeneracijski učinak treba razlikovati od stvarnog transgeneracijskog utjecaja do kojeg dolazi kad generacija u kojoj su učinci vidljivi nije niti bila izložena inicijalnom čimbeniku okoliša. Po ženskoj liniji bi to zapravo bila F3 generacija a po muškoj F2 i iduće generacije (slika 1).
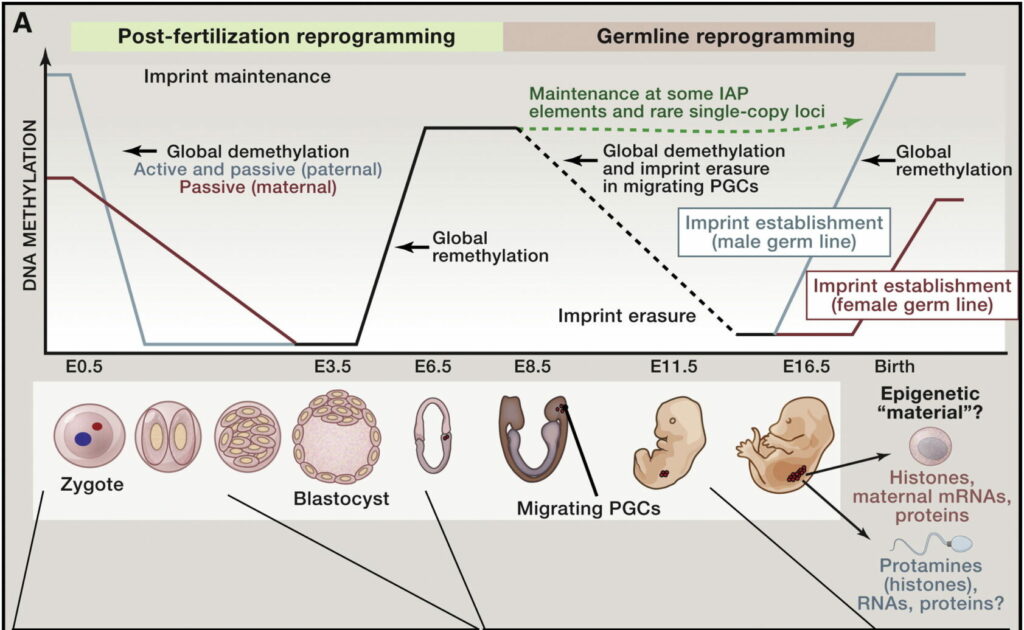
Na ovom mjestu treba spomenuti i epigenetičko reprogramiranje spolnih stanica odnosno činjenicu da se gotovo sve epigenetske oznake koje su postojale u jajnoj stanici i spermiju nakon oplodnje „brišu“. Do toga dolazi kako bi se omogućila totipotentnost oplođene jajne stanice (zigote) odnosno mogućnost da iz nje nastanu sve stanice novog organizma. Daljnjim diobama zigote nastaje rani embrij pri čemu dolazi do daljnjeg brisanja starih te uspostave novih obrazaca epigenetskih oznaka koje će biti karakteristične za tu jedinku i njene spolne stanice. Ovaj fenomen brisanja i uspostave novih epigenetskih oznaka (najčešće kroz metilaciju molekule DNA) naziva se još i genomskim upisivanjem (engl. genomic imprinting) te se smatra drugom barijerom prijenosu epigenetičke informacije na potomstvo.
Kad ovog mehanizma reprogramiranja ne bi bilo, to bi značilo da se sve epigenetske oznake u spolnim stanicama koje su naslijeđene ili stečene tijekom života prenose dalje na iduću generaciju. To bi moglo biti izrazito opasno jer bi se tako mogli naslijediti i neki nepovoljni učinci, primjerice poznato je da su epigenetski obrasci izmjenjeni kod nekih bolesti kao i tumora pa nasljeđivanje takvih stečenih „loših“ obrazaca ne bi bilo korisno za organizam. No ipak, postoje ideje kako se ipak ne gube sve epigenetske informacije te kako bi molekule koje prenose takvu vrstu informacije kroz generacije mogle biti male molekule RNA
jer preliminarne studije na nižim organizmima ukazuju na tu mogućnost (slika 2). Detalji tog procesa još nisu u potpunosti razjašnjeni na molekularnoj razini a nisu niti dokazani kod sisavaca, koji su nam naravno antropocentrično najzanimljiviji.
Mogućnost da se epigenetske oznake nasljeđuju nalazi svoje utemeljenje i u biljnom svijetu gdje je moguće iz somatske stanice biljke uzgojiti cijelu biljku – to je ono što bi se popularno zvalo „pelcanje“ a zapravo je riječ o kloniranju. Dakle kod biljaka nastanak nove jedinke ne mora uključivati spolne stanice (tučak-prašnici i plod kao rezultat) već se biljke mogu razmnožavati i nespolno što je zapravo kloniranje. Nasljeđivanje stečenih svojstava putem epigenetskih oznaka pokazano je i u životinjskom svijetu na nižim evolucijskim granama primjerice kod nematoda (oblića), no tako nešto se još uvijek smatra nemogućim kod sisavaca. Povremeno se može pojaviti poneka studija transgeneracijskog epigenetičkog nasljeđivanja, no upitno je je li se uistinu radi o prethodno opisanom transgeneracijskom ili intergeneracijskom učinku. Također se rijetko utvrđuje jesu li to uistinu epigenetičke promjene ili se ipak radi o mutacijama molekule DNA. Ako malo bolje razmislimo, intergeneracijski učinci su očito mogući u sisavaca, no jesu li mogući prijenosi epigenetskih obrazaca na generaciju potomaka koja nije bila prisutna u trenutku kada je pretpostavljeni snažni vanjski čimbenik poput stresa ili gladi djelovao? Takav slučaj odgovarao bi prijenosu na generaciju koja nije bila izložena tom agensu (prijenos s generacije F0 na F3 u ženskoj ili F2 u muškoj liniji, slika 1) a svejedno je naslijedila epigenetske obrasce prethodne generacije F0 koja je tome bila izložena.
U proteklih deset-petnaest godina su prilično odjeknule studije u kojima se tvrdilo kako unuci osoba koje su preživjele veliki i dugotrajni stres poput gladi ili boravka u koncentracijskom logoru mogu naslijediti neke karakteristike koje su posljedica upravo transgeneracijskog epigenetičkog nasljeđivanja. Radilo se o studijama koje su istraživale nasljeđivanje metaboličkog rizika u potomaka osoba koje su bile izložene takvoj vrsti stresa. Drastične promjene prehrane roditelja poput gladovanja i izloženost djeteta in utero takvim stresorima mogu rezultirati povišenim metaboličkim rizikom potomstva (Hales-Barker teorija, 2013). Jasno je da izloženost trudnica različitim štetnim agensima poput najočitijih nikotina i alkohola dokumentirano može utjecati na dijete no ovdje je riječ o intergeneracijskom epigenetičkom nasljeđivanju.

Kao jedan od potencijalnih primjera transgeneracijskog epigenetičkog utjecaja i nasljeđivanja često se koristi period gladi u Nizozemskoj pred kraj Drugog svjetskog rata. Utvrđeno je naime da su osobe koje su in utero bile izložene lošijoj prehrani majke imale slabiju toleranciju na glukozu u odnosu na osobe čije majke nisu prošle taj period gladi. Ovakav fenomen bio bi primjer utjecaja okoliša na razvoj embrija kako bi se potomstvo pripremilo na buduće loše uvjete i nedostatak hrane tijekom života. No ono što je bilo zapanjujuće jest da su i unuci žena koje su tijekom trudnoće prošle kroz period gladi imali povišen neonatalni adipozitet, dok su unuci muškaraca koji su prošli kroz period gladi in utero bili više skloni pretilosti u odnosu na osobe čiji preci nisu prošli takve stresove. Zanimljivo je također da kod ovih muškaraca nisu potvrđeni učinci pothranjenosti njihovih baka po majčinoj liniji. Iako medijski predstavljena kao primjer transgeneracijskog epigenetičkog nasljeđivanja ona to ustvari nije. Pokazalo se kako je kod osoba izloženih gladi in utero uistinu bila snižena razina metilacije DNA gena IGF2 u uzorcima pune krvi. No ovdje je i dalje teško razlučiti da se ne radi o učincima izloženosti koji se prenose direktno iz generacije u generaciju a ne transgeneracijski.
Osmišljavanje istraživanja transgeneracijskog epigenetičkog nasljeđivanja izrazito je kompleksno. Kako bi se izbjegli učinci in utero izloženosti ženskih potomaka i potencijal za intergeneracijsko nasljeđivanje, novija su istraživanja fenomena transgeneracijskog epigenetskog nasljeđivanja osmišljena tako da uključuju mušku liniju. Tako su primjerice miševi čija je hrana sadržavala niske količine proteina prenosili fenotip visokog kolesterola na svoje potomke po muškoj liniji. Pretpostavlja se da do toga dolazi zbog promjena u spermi, nije uočeno da bi se radilo o značajnije izmijenjenom obrascu metilacije DNA kao najčešćem obliku epigenetske promjene pa se pretpostavlja da se radi o promjenama na razini kromatina ili malih molekula RNA.
Jedan od možda najmanje upitnih primjera intergeneracijskog epigenetičkog nasljeđivanja, koje se potencijalno proteže i u transgeneracijsko, jest fetalna izloženost povišenim razinama tiroidnog hormona. Ovaj je fenomen uočen kod stanovnika Azora koji nose mutaciju u beta-receptoru za tiroidni hormon (THRbeta). Osobe koje su heterozigoti za tu mutaciju imaju povišene razine tiroidnog hormona u serumu koje ipak ne izazivaju nikakve kliničke manifestacije u smislu bolesti. Kod majki koje su heterozigoti za ovu mutaciju, djeca koja ju ne naslijede također su tijekom razvoja u maternici izložena povišenoj razini tiroidnog hormona. Dakle djeca nemaju mutaciju ali su in utero bila izložena povišenim razinama ovog hormona i pokazuju nižu porođajnu težinu. U odrasloj dobi također imaju slabiji odgovor TSH na egzogeno dodani tiroidni hormon što pokazuje da su naslijedili tu smanjenu senzitivnost. Djeca muškaraca F1 generacije, dakle onih koji su bili izloženi povišenom tiroidnom hormonu u maternici, također pokazuju tu smanjenu senzitivnost, dok djeca majki F1 generacije ne pokazuju taj fenomen. Isti obrazac se uočava u F3 generaciji kod osoba čije su prabake bile nositelji mutacije ili su im djedovi bili izloženi povišenim razinama tiroidnog hormona in utero. Ovaj primjer pokazuje kako se određeni fenotip, a to je senzitivnost na tiroidni hormon nasljeđuje neovisno od mutacije u molekuli DNA te je ujedno i jedan od „najčišćih“ primjera kako se neki fenotip može naslijediti ovisno o izloženosti pretka nekom stresoru – u ovom slučaju povišenim razinama tiroidnog hormona in utero. Ipak, niti u ovom primjeru ne mogu se isključiti neki drugi čimbenici koji su doveli do tog fenotipa.

Još jedan od poznatijih primjera koji se predstavlja kao potencijalno transgeneracijsko epigenetičko nasljeđivanje jest traumatsko iskustvo roditelja koje se prenosi na potomstvo kroz generacije za što se koristi primjer Holokausta. Već u startu je jasno da je vrlo teško u nečijem životu kao jedini značajni čimbenik izdvojiti neki traumatski događaj pa makar i produženog tipa te isključiti primjerice jednako tako moguć scenario da su roditelji koji su proživjeli traumu na drugačiji način odgajali djecu te tako utjecali na njihov fenotip. U studiji je pokazano kako su odrasli potomci osoba koje su preživjele Holokaust imale smanjene razine metilacije DNA gena FKBP5 u krvi. Radi se o genu koji kodira za jedan od bitnih regulatora osjetljivosti glukokortikoidnog receptora te je stoga bitan u odgovoru na stres no mogu li se takve promjene prenijeti i u iduću generaciju koja nije bila direktno izložena stresoru, nije moguće još uvijek sa sigurnošću utvrditi. Radilo se također o studiji s malim brojem ispitanika te je analiziran i manji set gena što je dodatna otegotna okolnost za validiranje ovih rezultata.
Zaključno možemo reći kako je i dalje biološki konsenzus da je Mendelov model nasljeđivanja u kojem se fenotip može naslijediti jedino prijenosom zapisa u molekuli DNA kroz generacije još uvijek jedini dokazani prijenos genetičke informacije u viših sisavaca. No, pojava novijih studija na modelnim organizmima, mahom nižeg evolucijskog stadija, pokazuje kako se neki fenotipovi možda mogu naslijediti preko spolnih stanica a da se ne radi o izmjenama zapisa u molekuli DNA.
Takav ne-genetički oblik nasljeđivanja temelji se na ideji da se neke druge molekule, osim molekule DNA, u spolnim stanicama mogu nasljeđivati te modificirati fenotip potomstva čak i kroz nekoliko generacija. Najčešće su takve promjene rezultat nekih ekstremnih izlaganja čimbenicima poput gladi ili nekih kemijskih agenasa koje utječu upravo na te „druge“ molekule (one koje nisu DNA, moguće da se radi o malim molekulama RNA) u spolnim stanicama te tako taj učinak prenose na potomstvo.
Postoji li uistinu ovakav tip nasljeđivanja kod ljudi je još predmet diskusije. Dok se u većem broju pravilno osmišljenih epidemioloških studija i studija na životinjskim modelima ne dokaže kako se uistinu radi o pravom transgeneracijskom epigenetičkom nasljeđivanju a ne o intergeneracijskom nasljeđivanju te isključi mogućnost drugih učinaka poput loše prehrane nakon rođenja koja je također mogla utjecati na epidemiološke studije koje su navedene kao primjeri, tema ostaje i dalje kontroverzna. Hoće li ih ikada i biti u ljudskoj populaciji također je upitno, jer bi takve studije morale biti dugotrajne budući da mogu proći desetljeća između inicijalne izloženosti predaka i potencijalne manifestacije u fenotipu potomaka. Dodatno, mnogi fenotipovi koji bi bili zahvaćeni ovakvim utjecajem su zapravo kompleksna svojstva poput primjerice sklonosti obolijevanja od nekih bolesti te su po svojoj prirodi podložna nizu utjecaja okoline koji se akumuliraju kroz život i rezultat su životnog stila pojedinca. Ono što bi se takvim studijama moralo nedvosmisleno utvrditi jest da je primjerice neka osoba oboljela od dijabetesa jer je njezina baka bila izložena stresu gladovanja i prenijela tu sklonost na potomstvo a ne zbog vlastitog životnog stila osobe. Dok se ne utvrdi molekularna osnova ovih mehanizama na životinjskim modelima te identificiraju molekule s istom ili sličnom funkcijom u ljudi ove će opservacijske studije ostati samo u domeni špekulacija.
1) Sharma U. Paternal contributions to offspring health: role of sperm small RNAs in intergenerational transmission of epigenetic information. Front Cell Dev Biol. 2019;7:215.
2) Senaldi L, Smith-Raska M. Evidence for germline non-genetic inheritance of human phenotypes and diseases. Clin Epigenetics. 2020;12(1):136
3) Cheuquemán C, Maldonado R. Non-coding RNAs and chromatin: key epigenetic factors from spermatogenesis to transgenerational inheritance. Biol Res. 2021;54(1):41.
4) Arzate-Mejía RG, Mansuy IM. Epigenetic Inheritance: Impact for Biology and Society-recent progress, current questions and future challenges. Environ Epigenet. 2022;8(1):dvac021
5) Maximilian Fitz-James, Giacomo Cavalli. Molecular mechanisms of transgenerational epigenetic inheritance.Nature Reviews Genetics, 2022, 23 (6), pp.325-341.
6) Fallet M, Blanc M, Di Criscio M, Antczak P, Engwall M, Guerrero Bosagna C, Rüegg J, Keiter SH. Present and future challenges for the investigation of transgenerational epigenetic inheritance. Environ Int. 2023;172:107776.
 5.00
(4)
5.00
(4)
25. ožujka 2026. / U fokusu

24. ožujka 2026. / U fokusu

17. ožujka 2026. / U fokusu

16. ožujka 2026. / U fokusu

27. ožujka 2026. / Perspektive Video

27. ožujka 2026. / Perspektive Publikacije

23. ožujka 2026. / Klub Batina Perspektive Rasprave

17. ožujka 2026. / Klub Batina Rasprave

{kind=link}
{kind=link}
_1a.jpg){kind=link}